
El águila de Haast Hieraaetus moorei; nuevas revelaciones sobre una especie extinta
Las primeras evidencias de esta especie de rapaz se obtuvieron en la década de 1870, en la isla sur de Nueva Zelanda, actualmente se considera que pertenece al género Hieraaetus, sobre su dieta se considera que estaba especializado en la caza de especies del género Dinornis, una de sus especies llegaba a pesar hasta 240 kg. El águila de haast hasta el momento de su extinción y en comparación con las especies actuales es la más grande, las hembras llegaban a pesar hasta los 15 kg y con una envergadura de hasta 3 metros. Con la llegada de los maoríes a Nueva Zelanda, las reglas del juego cambiaron, la extinción de las especies de Dinornis, provoco un efecto cascada y más tarde se cree que como consecuencia de la extinción de sus principales presas, el águila de haast llego a extinguirse, una nueva investigación describe sus hábitos alimenticios y la sitúan más próxima a las águilas cazadores que a los grandes carroñeros.
Ignacio García Dios
Text in English after Spanish
A finales del mes de octubre de 2021, vi un dibujo de uno de los mejores artistas ilustradores de fauna extinta como es Gabriel Ugueto. Sus dibujos sobre dinosaurios y otra fauna extinta son insuperables. Dispone de página web (www.gabrielugueto.com), donde poder disfrutar de ellos. En ese dibujo se podía leer:
Family of Haast’s Eagles (Hieraaetus moorei), one of the largest birds of prey ever to exist, sadly extinct since around the 1300s. The eagle’s main prey were all the species of Moa that existed in before humans arrived in NZ sometime around 1280. When all Moa species went extinct, so did their main predator. This illustration appears in book 5 (Tyrannosaurus rex) of the Extinct series written by professor Ben Garrod – Presenter.
Traduciendo ese texto, el autor nos estaría mostrando el dibujo de una familia de águila de Haast (Hieraaetus moorei), la cual se estima que se extinguió alrededor de 1300. Se encontraba en Nueva Zelanda y estaba especializada en la captura y consumo de especies del género Dinornis (moas). A finales del siglo XIII, como consecuencia de la llegada del ser humano a Nueva Zelanda, las moas se convirtieron en el objetivo de las partidas de caza de los maoríes, extinguiéndola rápidamente, lo cual provoco también la extinción de su principal depredador, el águila de Haast.

Es Julius von Haast, director del Museo de Canterbury, realizando una excavación de moas en la ciénaga de Glenmark (situada en la Isla Sur de Nueva Zelanda), el que localiza los primeros restos óseo del águila de Haast, la cual nombró como Harpagornis moorei, haciendo honor al propietario de la zona donde estaban realizando las excavaciones (George Moore).
Desde su descubrimiento ha sido clasificada en diversos géneros en Harpagornis y Aquila, actualmente se le asigna al género Hieraaetus. Este género esta formado por 5 especies más una extinta. Según el estudio realizado por Lerner et al. (2017), Phylogeny and new taxonomy of the Booted Eagles, basado en el análisis molecular de siete loci, se establece que en 3 de los 4 análisis realizados, el género Hieraaetuses monofilético. Existe un antecesor común a las seis especies. A través de otro antecesor común, divergen alrededor de 1 m.a. las especies Hieraaetus weiskei (Nueva Guinea e islas Molucas septentrionales), Hieraaetus morphnoides (Australia) y Hieraaetus moorei (†; Nueva Zelanda). Individuos de la subespecie de Hieraaetus pennatus pennatus, podrían haber llegado a esas zonas a través de sus procesos de migración hacia el sur (Gjershaug et al., 2009).
Así se describe esta especie en García Dios (2017), águila de haast (extinguida), Aquila moorei (Haast, 1872). Su extinción se asocia al momento en que se extingue su presa principal (moas) por parte de los Maoríes y por la destrucción de su hábitat. Solo se ha constatado su existencia en la isla Sur de Nueva Zelanda. Fue el águila de mayor tamaño conocida. Su presa principal eran las moas, aves gigantes no voladoras que eran incapaces de defenderse de la fuerza de choque y la velocidad de estas águilas, las cuales se considera que podían alcanzar una velocidad de 80 km/h. El gran tamaño del águila puede haber sido una respuesta evolutiva al tamaño de sus presas, ya que habría sido sustancialmente menor cuando arribó a la isla, para luego crecer sustancialmente a lo largo del tiempo debido a la carencia de competencia cuando su principal fuente alimenticia, las moas. Después de realizar diversos análisis de ADN, el águila de haast se podría reclasificar como Hieraaetus moorei al haber divergido de especies como H. pennatus y H. morphnoides entre 700.000-1,8 m.a. Presentaban RSD en tamaño a favor de las hembras. Peso estimado entre 10-15 kg en las hembras; machos sobre 9-12 kg.; envergadura relativamente corta entre 2,6-3 m. Alas cortas adaptadas a cazar entre densos matorrales y bosques. Tarsos fósiles presentan dimensiones sobre 22,7-24,9 cm. Cola muy larga hasta 50 cm. Longitud corporal de 1,4 m en hembras y una altura en posición erguida de 90 cm.

Fotografía / Photography: Otago Museum (Dunedin, New Zealand).
Los moas eran aves muy grandes del género Dinornis. De hecho, su nombre científico se podría traducir por “pájaros dinosaurio«. En un nuevo estudio publicado sobre el águila de Haast en Proceedings of the Royal Society B: Biological Sciences. 288 (1964) en diciembre de 2021 por A. H. van Heteren, S. Wroe, L. R. Tsang, D. R. Mitchell, P. Ross, J. A. Ledogar, M. R. G. Attard, D. Sustaita, P. Clausen, R. P. Scofield y G. Sansalone New Zealand’s extinct giant raptor (Hieraaetus moorei) killed like an eagle, ate like a condor, nos muestran más datos sobre estas especies de aves gigantes.
Lo primero que me ha sorprendido en esta publicación, es el peso medio de un moa adulto el cual se estima sobre 200 kg. En otras publicaciones, se están recogiendo pesos de hembras de alrededor de 240 kg, y de machos de 85 kg. Cuando llegaron los moas a Nueva Zelanda, no presentaban este marcado dimorfismo sexual, ni tampoco estas grandes dimensiones. El por qué fueron incrementando su tamaño, es como consecuencia de la ausencia en las islas de algún mamífero de gran tamaño. Esto permitió que los moas fuesen aumentando su tamaño hasta convertirse en los herbívoros de mayor tamaño de su ecosistema. En cuanto al elevado dimorfismo, puede explicarse que las hembras más grandes de moas, pondrían más huevos, siendo seleccionadas por los machos. Parece ser que este proceso fue bastante rápido como se puede ver por el registro fósil.
Llegó un momento en que los moas terminaron prescindiendo de las alas, las cuales les resultaban completamente inútiles. No eran capaces de volar, y al mismo tiempo, no sufrían una gran presión de los depredadores presentes en las islas. Solo el águila de Haast era el único depredador que podía depredar sobre ellas. Con la llegada del ser humano, las reglas cambiaron. Al llegar los maoríes, y descubrir estas imponentes aves, comenzaron a cazarlas. Según los últimos estudios, en poco más de 100 años consiguieron acabar con todos los ejemplares de cada una de las nueve especies que formaban el género.
El águila de Haast, sería una especie entre un 30-40% más grande que la mayor de las águilas actuales, el Águila arpía, (Harpia harpyja). Como ya hemos hablado anteriormente, este águila procede de un ancestro común de las Hieraaetus, siendo su antecesor más próximo una de las águilas más pequeñas del mundo, otra rapaz australiana, el águila australiana (Hieraaetus morphnoides). Estamos ante un ejemplo extraordinario de evolución rápida en menos de 2 millones de años.
Los maoríes la llamaban Pouakai, Hokioi o Hakawai (viejo glotón). Según Sir George Gray, Gobernador de Nueva Zelanda en dos etapas (1845-1854 y 1861-1868), era un ave de color blanco y negro, con una cresta roja sobre su cabeza y las puntas de las alas teñidas de amarillo y verde. Los maoríes la respetaban y temían. Viejas leyendas transmitidas oralmente, hablan de raptos de seres humanos y en el país abundan las pinturas del depredador en rocas y cuevas.
Este pájaro, el hokioi, lo conocieron nuestros antepasados. Nosotros no la hemos visto, es un pájaro que ha desaparecido. Pero lo que decían nuestros antepasados era que era poderosa, un ave muy poderosa. Un halcón muy grande. Descansaba en la cima de las montañas, no descansaba en las llanuras. Cuando volaba nuestros antepasados lo veían pero no todos los días, porque moraba en las montañas. Era rojo, blanco y negro. Era un pájaro de plumas negras teñidas de amarillo y verde y un montón de plumas de color rojo en lo alto de la cabeza. Esta es la descripción que hace un maorí a Sir James Hector, geólogo refutado, en una reunión de naturalistas de la Royal Society of New Zealand el 14 de agosto de 1872.

Ahora, y tras una nueva y extensa investigación, los científicos creen que no se trata solo de una simple leyenda.
Sobre el águila de Haast, se ha escrito mucho sobre si era depredadora o carroñera. Por su estructura ósea se creía que basaba su dieta en el consumo de carroñas, la estructura de las fosas nasales, etc. Las representaciones de esta especie, siempre la muestran con una cabeza desplumada lo que haría pensar a una adaptación para acceder al interior de los cuerpos sobre los que se alimentaba. Por contra, las leyendas maoríes hablaban de un águila capaz de depredar sobre humanos, lo cual quedó reflejado en arte rupestre.

En los últimos estudios sobre el águila de haast, apoyan la teoría de la especie depredadora. En este último estudio, se describe a una especie capaz de abatir a sus presas en picados de hasta 80 km/h, así como el de disponer de unas estructuras faciales, que le permitieran introducir sus cabezas en el interior de las presas para alimentarse. Basándose en el sistema nervioso, entre otras características, se determina que la inervación bien desarrollada de las extremidades posteriores indica que sus garras eran lo suficientemente sensibles y poderosas para agarrar presas vivas, y carecía de algunas características visuales, olfativas y vestibulares que son comunes a muchos buitres. La hipótesis de la caza se ha visto respaldada por análisis de forma bidimensional y estudios cualitativos que han demostrado una similitud general en la forma del cuerpo y la morfología de las garras con las grandes águilas existentes.
En el estudios publicado de Heteren et al (2021), basado en el análisis comparativo de la forma y la fuerza biomecánica de neurocráneos, picos y garras en cinco aves carroñeras y depredadoras existentes, muestra un ave cazadora como las águilas, pero como ocurre con algunas especies actuales de otros géneros de águilas que se presumían como cazadoras (Margalida et al. 2017), también se alimentaba de carroñeras. El neurocráneo de H. mores tiene forma de buitre, mientras que su pico es de águila. El desempeño mecánico es más cercano al de las águilas existentes con bajo cargas de mordedura, pero es más cercano al del cóndor andino (Vultur gryphus) bajo cargas extrínsecas que simulan la captura y matanza de presas. Las garras, sin embargo, son parecidas a las de un águila e incluso para un ave de su tamaño, son capaces de soportar cargas extremadamente altas. Los resultados son consistentes con la proposición de que, a diferencia de las águilas vivas, las águilas de Haast habitualmente mataban presas más grandes que ellas mismos, lo cual implicaba que se alimentase durante varios días de una presa tan grande. Como eran las moas. El desacoplamiento de la relación entre el neurocráneo y la forma del pico puede haber estado relacionado con una evolución rápida.
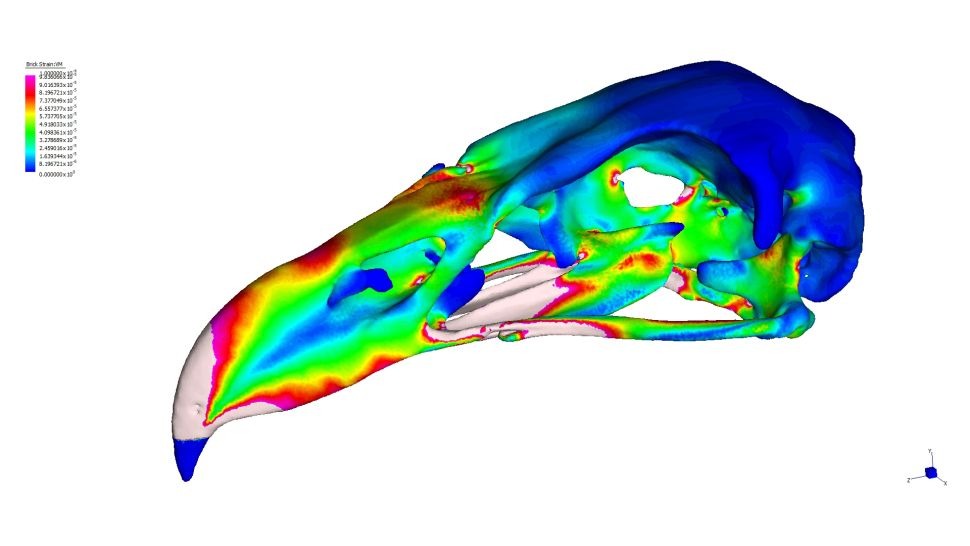
En esta investigación, además de H. moorei, se utilizaron muestras de buitre negro (Aegypius monachus), cóndor andino (Vultur gryphus), águila audaz (Aquila audax), águila australiana (Hieraaetus morphnoides) y milano silbador (Haliastur sphenurus). Se realizaron los siguientes análisis:
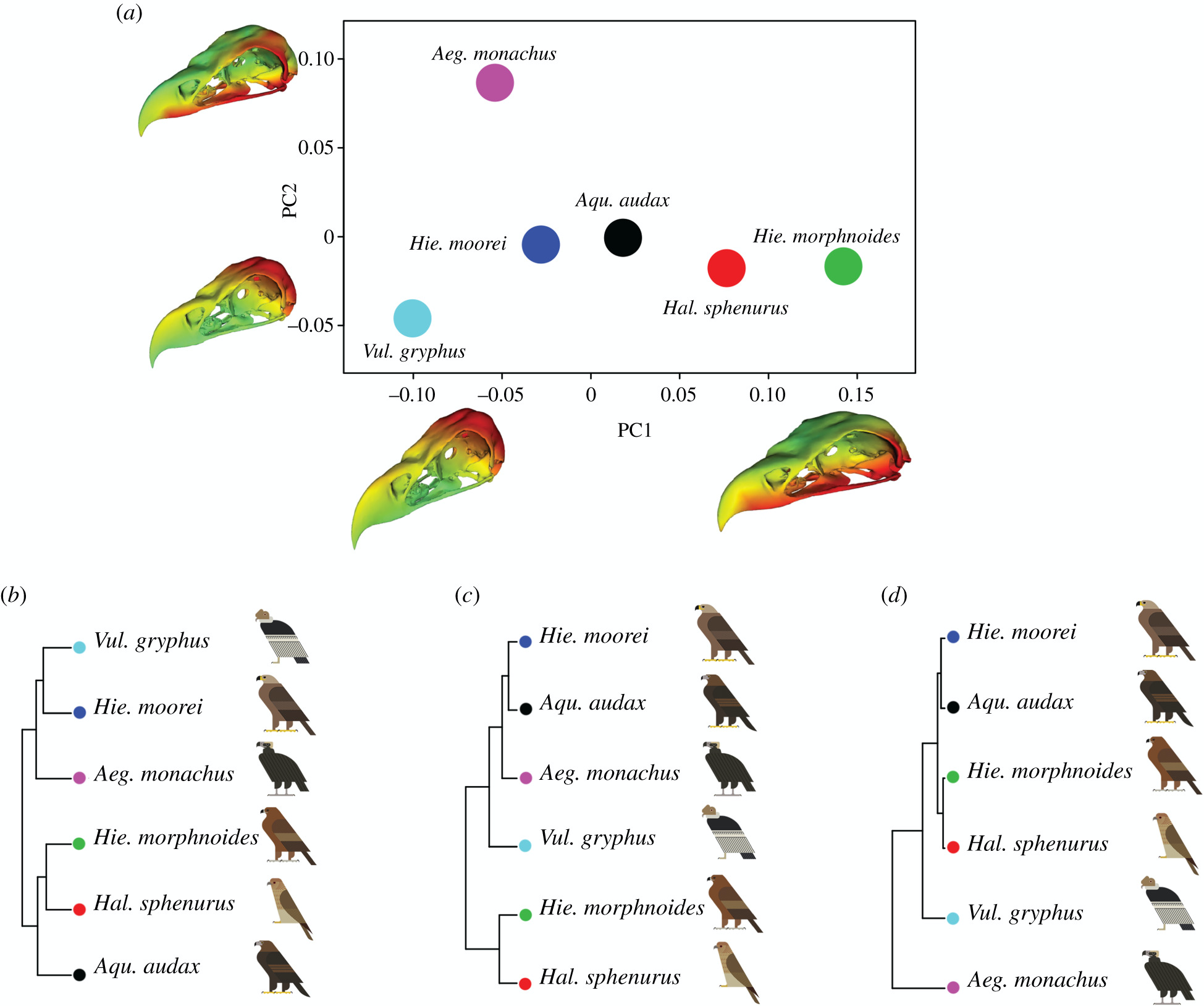
1. Análisis de la forma de los cráneos.- Gracias a la realización de análisis de componentes principales (PCA; PC1, PC2, PC3), se revelaron diferencias morfológicas entre las especies de aves rapaces según el modo de alimentación. El cráneo con un pico más corto y un neurocráneo más ancho y plano, serían las propias de las especies más pequeña (H. esfenuro y H. morphnoides). Las especies más grandes (A. audax, H. moorei y V. gryphus) mostraron un neurocráneo más estrecho y un pico más largo. Las dos especies de buitres y H. moorei tenían un neurocráneo más alto y estrecho, mientras que las otras águilas y el milano tenían un neurocráneo más ancho. Realizando un análisis UPGMA (método simple de agrupamiento jerárquico basado en matrices de disimilitud), realizado en el conjunto completo de variables de forma reveló similitudes cercanas entre H. moorei y ambos buitres. Sin embargo, los UPGMA realizados en PC1 y PC2 registrados por separado muestran que H. moorei se parecía más a A. audax. Este resultado para registros PC1 separados se explicó en gran medida por la similitud en la forma del pico entre estas dos especies de águilas grandes. UPGMA basado en PC2 destacó las diferencias entre los dos carroñeros existentes (V. gryphus y A. monachus), al menos en parte es explicada por la filogenia. UPGMA en PC3 mostró que H. moorei es más parecido a un buitre con respecto a la forma general del cráneo, en gran parte debido a las similitudes en la orientación del foramen magnum y el ancho craneal relativo.
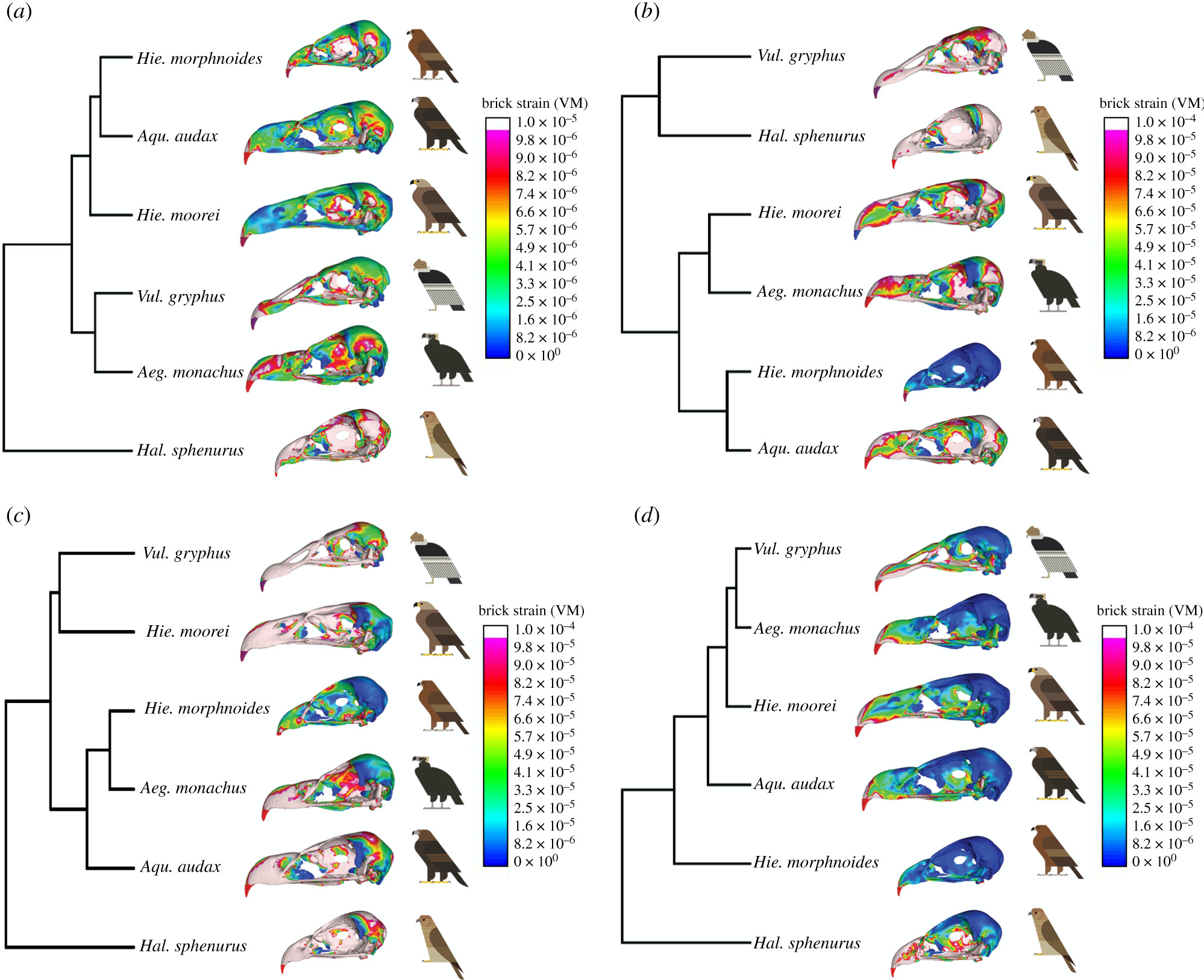
2. Análisis de elementos finitos de los cráneos.- En el caso de carga intrínseca (mordiente), la deformación media fue notablemente menor en H. moorei que en todos los demás taxones, aunque más cercano a A. monachus. Los valores de tensión fueron similares entre las especies restantes, excepto H. sphenurus en el que los valores de tensión fueron altos; Los resultados de UPGMA no fueron consistentes con estos valores medios, pero mostraron una gran similitud de H. moorei con A. audax y H. morphnoides. A diferencia de ambos buitres, pero como A. audax, H. moorei mostró tensiones extremadamente bajas en la mitad del pico.
Para la carga dorsoventral, la deformación media fue intermedia en H. moorei, cayendo entre el registrado para V. gryphus y A. monachus. Los valores para A. monachus también fueron intermedios y los valores para A. audax y H. morphnoides fueron muy bajos, especialmente para H. morphnoides. H. sphenurus presentó la tensión más alta. Estos resultados fueron consistentes con los de la UPGMA. El patrón de tensión en H. moorei era más parecido al de A. audax, con una banda de tensiones altas a lo largo de la caja craneana y una mancha de tensiones bajas en el lateral del pico. En la sacudida lateral, tensión media en el cráneo de H. moorei era relativamente alta y similar al de V. gryphus. Las dos especies de águilas existentes registraron registros bajos, aunque los más bajos fueron en A. monachus y los más altos en H. sphenurus. Bajo la simulación de retroceso, H. moorei mostró una tensión relativamente baja, comparable a la de V. gryphus y A. monachus. A. audax y H. morphnoides registraron registros más altos, pero nuevamente la más alta fue en H. esfenuro. Estos resultados también se reflejan en la UPGMA.
3. Análisis de la forma de las garras.- El análisis de la forma de las garras separó claramente a los buitres de las demás rapaces. Los buitres tenían una garra más corta y roma con una curvatura disminuida, y una superficie articular ungueal reducida, mientras que los taxones de rapaces se caracterizaban por un eje ungueal más curvo, más largo y puntiagudo, y una cara articular ungueal más grande y robusta. El análisis UPGMA realizado en todas las variables de forma de la garra, mostró que H. moorei era más como H. morphnoides.
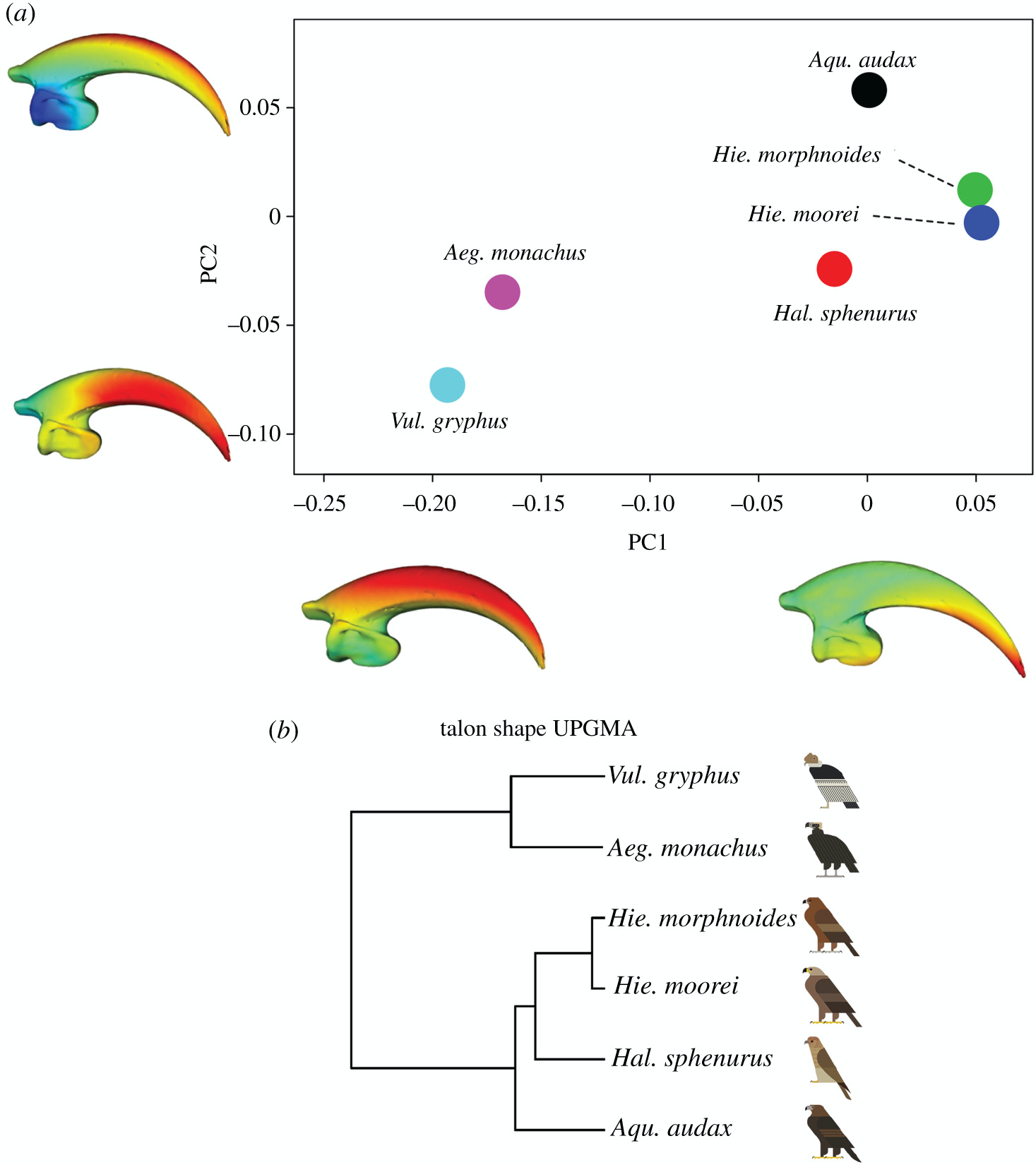

4. Análisis de elementos finitos de las garras.- Los datos de tensión media y el UPGMA de los resultados de tensión para las garras de las seis especies de rapaces, mostraron que H. moorei se parecía más a A. audax, el águila existente más grande en el análisis. Sin embargo, los datos de tensión media fueron considerablemente más bajos en H. moorei que en cualquiera de las otras rapaces. H. moorei mostró valores de tensión consistentemente más bajos en la región ventral de la curvatura interna, en la curvatura externa y en el tubérculo flexor en comparación con A. audax. H. morphnoides y H. esfenuro mostraron valores de deformación intermedios, mientras que las dos carroñeras mostraron la deformación más alta.
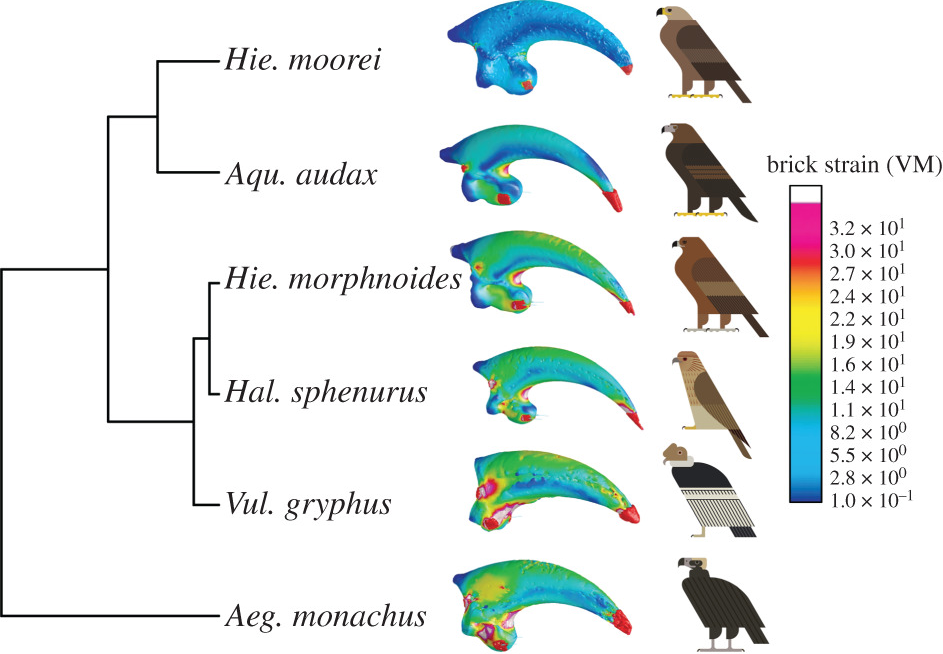
Una vez realizados las pruebas mecánicas, simulaciones y análisis, los autores presentan sus conclusiones, las cuales os las recojo a continuación:
Este trabajo, el cual no se ha basado como los anteriores en interpretaciones, comparaciones cualitativas, así como análisis limitados de formas bidimensionales, sino en análisis combinados los cuales han revelado un nuevo mosaico de características morfológicas y biomecánicas atribuibles al cráneo de H. moorei. En cuanto a la forma, el pico es de águila pero su neurocráneo es el más cercano al de los dos carroñeros, concretamente al del cóndor andino (V. gryphus) en particular. Esto contradice las interpretaciones basadas en análisis bidimensionales de que las similitudes más cercanas entre H. moorei y buitres eran evidentes en el pico.
Hieraaetus moorei también se caracterizó por una combinación de rasgos de águila y buitre con respecto al rendimiento mecánico del cráneo. Bajo la carga intrínseca, la deformación media en H. moorei fue considerablemente más baja que en cualquier otro taxón, pero más cercano al de las águilas. Esto sugiere una capacidad para soportar cargas generadas por una mordida particularmente poderosa para su tamaño. Deformaciones medias altas registradas en V. gryphus, H. esfenuro y H. moorei para la tracción dorsoventral sugiere que estas especies no están bien adaptadas para este comportamiento. De manera similar, la tensión media alta para H. moorei bajo la simulación de sacudidas laterales indica que esta especie estaba mal adaptada para realizar este comportamiento en relación con las águilas existentes y nuevamente estaba más cerca de V. gryphus que se alimenta de carroña. Curiosamente, el otro carroñero, el A. monachus, registra una tensión muy baja para la sacudida lateral. Bajo la carga de retroceso, H. moorey y los dos carroñeros se manejan mejor que los taxones restantes y el águila de Haast fue nuevamente la más cercana a V. gryphus, lo cual es consistente con las observaciones de alimentación y la anatomía funcional de los buitres.
La interpretación tanto del análisis de forma como de los resultados FEA de la garra muestran que H. moorei es inequívocamente parecido a un águila (mostrando similitudes con A. audax) y capaz de resistir cargas particularmente altas. Curiosamente, H. moorei mostró las similitudes morfológicas más cercanas con la Hieraaetus filogenéticamente más cercana: H. morphnoides. En el caso de H. moorei, el aumento en el tamaño de la garra probablemente refleja la adaptación a matar presas muy grandes en relación con la masa corporal del ave, pero en este caso, también podría deberse a la inercia filogenética.
En general, el pico de H. moorei es como un águila y su neurocráneo como un buitre, teniendo el mayor parecido con V. gryphus entre los dos carroñeros. Los resultados de las simulaciones FEA del comportamiento de alimentación muestran H. moorei funciona muy bien bajo la simulación de mordida, pero pobremente bajo cargas dorsoventrales y laterales. Bajo la carga de retroceso, H. moorei supera a las otras dos águilas y vuelve a estar más cerca de V. gryphus. Por lo tanto, con respecto a las cuatro simulaciones FEA, es más cercano a las águilas en la capacidad de dar un mordisco poderoso, pero más cercano a la especie que se alimenta de carroña como el cóndor andino. Su garra, por otro lado, es como un águila en todos los aspectos y es capaz de soportar grandes fuerzas, incluso para su gran masa corporal.
Entonces, ¿cómo interpretar esta combinación única de caracteres en H. moorei? Las garras son las principales herramientas de matanza de la mayoría de las aves rapaces, excepto los halcones; sin embargo, su pico puede usarse para terminar una captura de una presa. Proponemos que morder puede haber sido mas importante para matar a H. moorei de lo que es típico entre especies del género Aquila. Con respecto al FEA extrínseco, primero notamos que existe una clara diferencia entre los dos carroñeros conocidos incluidos en los análisis. A. monachus pudo soportar altas cargas laterales, mientras que H. moorei y V. gryphus se desenvuelven mal en este sentido. Esto es consistente con la clasificación de A. monachus como un desgarrador de carne, capaz de abrir la piel dura y consumir la carne de los cadáveres de grandes mamíferos, posiblemente con fuertes fuerzas laterales y poderosas mordidas. Aegypius monachus es un buitre grande y agresivo que domina a otras aves carroñeras cuando se producen peleas por un cadáver. Por el contrario, V. gryphus es un ‘tragador/engullidor’ que se centra en los órganos internos más blandos. Nuestros hallazgos de características más parecidas a buitres en los FEA extrínsecos, por lo tanto, son más consistentes con la proposición de que H. moorei puede haberse centrado preferentemente en las vísceras ricas en nutrientes de sus presas, mismo comportamiento que la mayoría de las aves rapaces que se alimentan de carroña.
Esta mezcla de morfologías y comportamientos mecánicos que recuerdan a los carroñeros obligados, así como a las águilas depredadoras, puede ser el resultado de una combinación única de presiones selectivas. Los buitres generalmente se alimentan de animales mucho más grandes que ellos, pero las águilas suelen matar y comer presas relativamente pequeñas, sin embargo también se pueden alimentar de carroñas de grandes presas. La evidencia tafonómica deja pocas dudas de que H. moorei mató y se alimentó de moas, incluido los más grandes del género Dinornis, que podía pesar más de 200 kg. Incluso la especie de moa más pequeña (moa arbusto pequeño, Anomalopteryx didiformis; aproximadamente 30 kg) era más grande que H. moorei. El águila de Haast probablemente habitó llanuras abiertas, al igual que muchas de sus especies presa. Evidentemente, su visión no era particularmente aguda, pero sin competencia, esto probablemente no estaba bajo una fuerte selección. H. moorei usó un vuelo de aleteo, lo que sugiere que el vuelo se produjo en distancias relativamente cortas. En conjunto, la evidencia sugiere que H. moorei podría haber descendido en picado desde árboles o acantilados a distancias cortas para atrapar y matar a su presa. Nuestros resultados son en gran parte consistentes con la proposición de que H. moorei habitualmente mataba presas particularmente grandes de una manera típicamente parecida a un águila usando sus poderosas garras, pero luego aplicaba técnicas de alimentación comunes a los buitres que comen de los cadáveres de especies mucho más grandes que ellos. La piel de las aves suele ser menos dura que la de los mamíferos, por lo que H. moorei fue capaz de abrir los cadáveres por sí mismo. Es notable que H. moorei estaría menos adaptada a comer tejidos duros, como tendones, que Aegypius, pero probablemente prefiera vísceras y músculos, como Vultur.
Nuestra propuesta que H. moorei prefería los órganos internos de sus presas, lo que lleva a la posibilidad de que, además de las volutas de las fosas nasales similares a las de los buitres y la mecánica de alimentación similar a la de los buitres, como la mayoría de los buitres, su cabeza y cuello no tenían plumas. Esta proposición también es consistente con una representación de arte rupestre maorí de lo que se cree que es un H. moorei, en la Cueva del Águila en la Estación Craigmore, Isla Sur. El ave rapaz tiene un cuerpo de color oscuro, pero la cabeza y el cuello permanecen sin color.

Haast’s eagle Hieraaetus moorei; new revelations about an extinct species
The first evidence of this species of raptor was obtained in the 1870s, in the South Island of New Zealand, it is currently considered to belong to the genus Hieraaetus, on its diet it is considered that it was specialized in hunting species of the genus Diornis, one of its species weighed up to 240 kg. The haast eagle until the moment of its extinction and compared to the current species is the largest, the females weighed up to 15 kg and with a wingspan of up to 3 meters. With the arrival of the Maori in New Zealand, the rules of the game changed, the extinction of the Diornis species caused a cascade effect and later it is believed that as a consequence of the extinction of its main prey, the haast eagle came to become extinct, new research describes its eating habits and places it closer to hunting eagles than to large scavengers.
At the end of October 2021, I saw a drawing by one of the best illustrating artists of extinct fauna, Gabriel Ugueto. His drawings of dinosaurs and other extinct fauna are second to none. He has a website (www.gabrielugueto.com), where you can enjoy them. In that drawing you could read:
Family of Haast’s Eagles (Hieraaetus moorei), one of the largest birds of prey ever to exist, sadly extinct since around the 1300s. The eagle’s main prey were all the species of Moa that existed in before humans arrived in NZ sometime around 1280. When all Moa species went extinct, so did their main predator. This illustration appears in book 5 (Tyrannosaurus rex) of the Extinct series written by professor Ben Garrod – Presenter.
Translating that text, the author would be showing us the drawing of a family of Haast’s eagle (Hieraaetus moorei), which is estimated to have become extinct around 1300. It was found in New Zealand and specialized in the capture and consumption of species of the genus Dinornis (moas ). At the end of the 13th century, as a consequence of the arrival of human beings in New Zealand, the moas became the target of the Maori hunting parties, quickly extinguishing them, which also caused the extinction of their main predator, the Haast eagle.
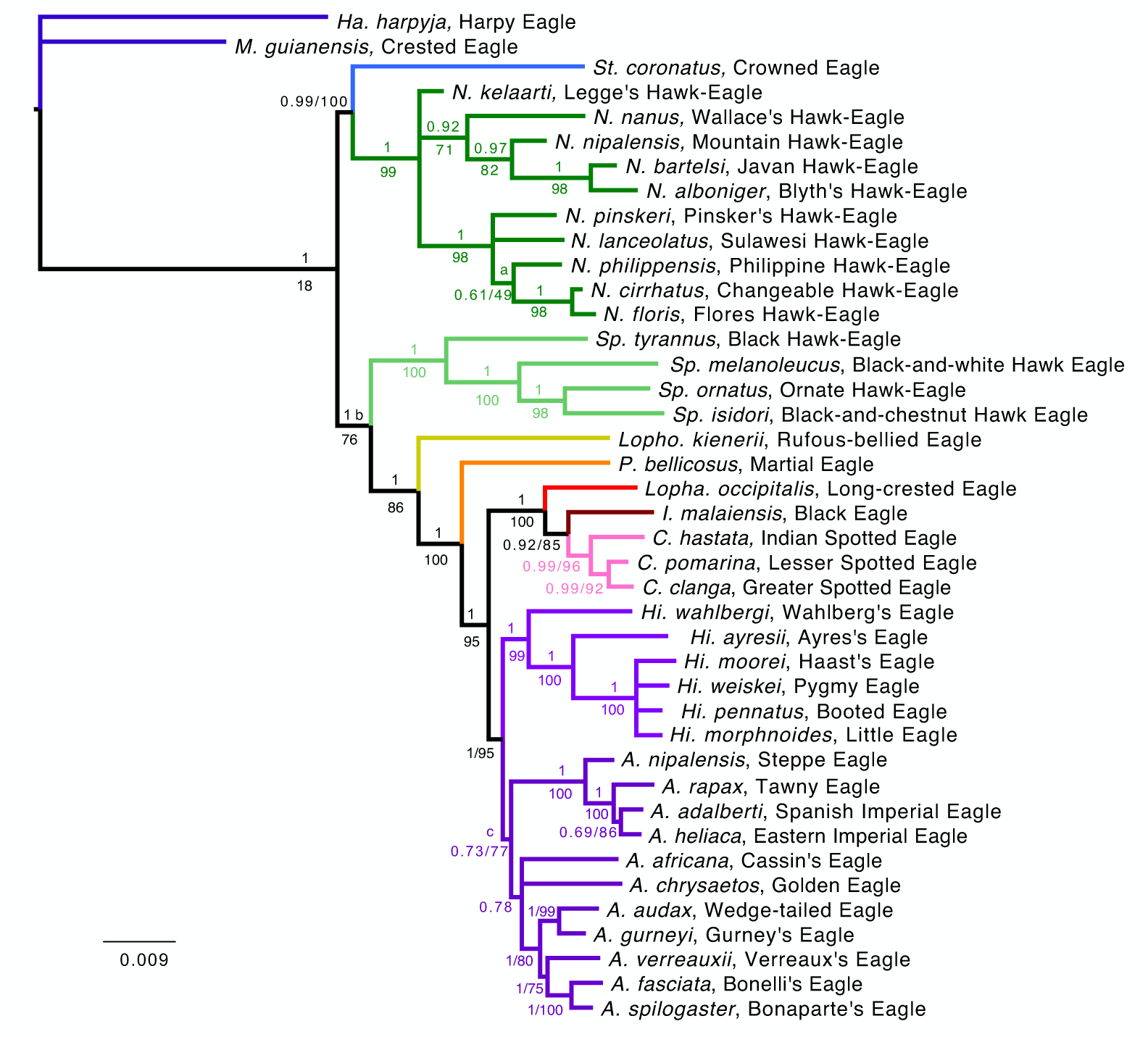
Phylogenetic tree based on the molecular study of seven loci published by Lerner et al. (2017)
It is Julius von Haast, director of the Canterbury Museum, excavating moas in the Glenmark swamp (located in the South Island of New Zealand), who locates the first bone remains of Haast’s eagle, which he named Harpagornis moorei , honoring the owner of the area where the excavations were being carried out (George Moore).
Since its discovery it has been classified in various genera in Harpagornis and Aquila, currently it is assigned to the genus Hieraaetus. This genus is made up of 5 species plus one extinct. According to
the study conducted by Lerner et al. (2017), Phylogeny and new taxonomy of the Booted Eagles, based on the molecular analysis of seven loci, it is established that in 3 of the 4 analyzes carried out, the genus Hieraaetus is monophyletic. There is a common ancestor to all six species. Through another common ancestor, they diverge about 1 m.y. the species Hieraaetus weiskei (New Guinea and northern Moluccas), Hieraaetus morphnoides (Australia), and Hieraaetus moorei (†; New Zealand). Individuals of the Hieraaetus pennatus pennatus subspecies could have reached these areas through their southward migration processes (Gjershaug et al., 2009).
Haast’s eagle (extinct), Aquila moorei (Haast, 1872). Its extinction is associated with the moment in which its main prey (Moas) is extinguished by the Maori and the destruction of its habitat. Its existence has only been confirmed in the South Island of New Zealand. It was the largest known eagle. Their main prey was the Moas, giant flightless birds that were unable to defend themselves against the shock force and speed of these eagles, which are considered to be able to reach a speed of 80 km/h. The large size of the eagle may have been an evolutionary response to the size of its prey, as it would have been substantially smaller when it arrived on the island, only to grow substantially over time due to lack of competition when its main food source, the Moas. After performing various DNA analyses, the haast’s eagle could be reclassified as Hieraaetus moorei, having diverged from species such as H. pennatus and H. morphnoides between 700,000-1.8 m.a. They presented RSD in size in favor of females. Estimated weight between 10-15 kg in females; males about 9-12 kg.; relatively short wingspan between 2.6-3 m. Short wings adapted to hunting in dense thickets and forests. Fossil tarsi have dimensions of about 22.7-24.9 cm. Very long tail up to 50 cm. Body length of 1.4 m in females and a height in an upright position of 90 cm.
Moas were very large birds of the genus Dinornis. In fact, its scientific name could be translated as «dinosaur birds». In a new study published on Haast’s eagle in Proceedings of the Royal Society B: Biological Sciences. 288 (1964) in December 2021 by AH van Heteren, S Wroe, LR Tsang, DR Mitchell, P. Ross, JA Ledogar, MRG Attard, D. Sustaita, P. Clausen, RP Scofield, and G. Sansalone New Zealand’s extinct giant raptor (Hieraaetus moorei) killed like an eagle, ate like a condor, show us more data about these species of giant birds.
The first thing that has surprised me in this publication is the average weight of an adult Moa, which is estimated to be over 200 kg. In other publications, weights of females of around 240 kg and males of 85 kg are being collected. When the Moas arrived in New Zealand, they did not present this marked sexual dimorphism, nor these large dimensions. The reason why they were increasing in size is as a consequence of the absence of any large mammal on the islands. This allowed the moas to increase in size until they became the largest herbivores in their ecosystem. Regarding the high dimorphism, it can be explained that the largest female Moas would lay more eggs, being selected by the males. It appears that this process was quite rapid as can be seen from the fossil record.
There came a time when the moas ended up doing without their wings, which were completely useless to them. They were not capable of flight, and at the same time, they did not suffer great pressure from the predators present on the islands. Only Haast’s eagle was the only predator that could prey on them. With the arrival of the human being, the rules changed. When the Maori arrived and discovered these imposing birds, they began to hunt them. According to the latest studies, in just over 100 years they managed to wipe out all the specimens of each of the nine species that made up the genus.
Haast’s eagle, would be a species between 30-40% larger than the largest of the current eagles, the Harpy Eagle, (Harpia harpyja). As we have already mentioned, this eagle comes from a common ancestor of the Hieraaetus, its closest ancestor being one of the smallest eagles in the world, another Australian bird of prey, the Little Eagle (Hieraaetus morphnoides). This is an extraordinary example of rapid evolution in less than 2 million years.
The Maori called it Pouakai, Hokioi or Hakawai (old glutton). According to Sir George Gray, Governor of New Zealand in two stages (1845-1854 and 1861-1868), it was a black and white bird, with a red crest on its head and the tips of its wings dyed yellow and green. The Maori respected and feared her. Old orally transmitted legends speak of abductions of human beings and in the country there are many paintings of the predator on rocks and caves.
This bird, the hokioi, was known to our ancestors. We have not seen it, it is a bird that has disappeared. But what our ancestors said was that it was powerful, a very powerful bird. A very large falcon. He rested on top of the mountains, he did not rest on the plains. When it flew, our ancestors saw it, but not every day, because it lived in the mountains. It was red, white and black. It was a bird with black feathers tinged with yellow and green and a bunch of red feathers on top of its head. This is the description made by a Maori to Sir James Hector, a refuted geologist, at a meeting of naturalists of the Royal Society of New Zealand on August 14, 1872.

Now, after extensive new research, scientists believe that it is not just a simple legend.
About Haast’s eagle, much has been written about whether it was a predator or a scavenger. Due to its bone structure, it was believed that it based its diet on the consumption of carrion, the structure of the nostrils, etc. The representations of this species always show it with a plucked head, which would suggest an adaptation to access the interior of the bodies on which it fed. In contrast, Maori legends spoke of an eagle capable of preying on humans, which was reflected in rock art.
In the latest studies on the haast eagle, they support the theory of the predatory species. In this latest study, a species capable of taking down its prey in dives of up to 80 km/h is described, as well as having facial structures that allow it to insert its heads inside the prey to feed. Based on the nervous system, among other characteristics, it is determined that the well-developed innervation of the hind limbs indicates that its claws were sensitive and powerful enough to grasp live prey, and it lacked some visual, olfactory, and vestibular characteristics that are common to many vultures. The hunting hypothesis has been supported by two-dimensional shape analysis and qualitative studies that have shown a general similarity in body shape and talon morphology to extant large eagles.
In the published study by Heteren et al (2021), based on the comparative analysis of the shape and biomechanical strength of neurocrania, beaks and claws in five extant scavengers and predators, it shows a hunting bird like eagles, but as it happens with some current species of other eagle genera that were presumed to be hunters (Margalida et al. 2017), also fed on scavengers. The neurocranium of H. moorei is shaped like a vulture, while its beak is like an eagle’s. Mechanical performance is closest to that of extant eagles under bite loads, but is closest to that of the Andean condor (Vultur gryphus) under extrinsic loads that simulate prey capture and killing. The claws, however, are eagle-like and even for a bird of its size, they are capable of supporting extremely high loads. The results are consistent with the proposition that, unlike living eagles, Haast’s eagles routinely killed prey larger than themselves, which meant feeding for several days on such large prey. How were the moas? The decoupling of the relationship between neurocranium and beak shape may have been related to rapid evolution.
In this research, in addition to H. moorei, samples of Cinereous Vulture (Aegypius monachus), Andean condor (Vultur gryphus), Wedge-tailed Eagle (Aquila audax), little eagle (Hieraaetus morphnoides), and whistling kite (Haliastur sphenurus) were used. The following analyzes were performed:
1. Analysis of the shape of the skulls.- Thanks to principal component analysis (PCA; PC1, PC2, PC3), morphological differences were revealed between the species of birds of prey according to the mode of feeding. The skull with a shorter beak and a broader and flatter neurocranium would be typical of the smaller species (H. sfenuro and H. morphnoides). The largest species (A. audax, H. moorei, and V. gryphus) showed a narrower braincase and longer bill. The two species of vultures and H. moorei had a taller and narrower braincase, while the other eagles and the kite had a broader braincase. Performing a UPGMA (Simple Hierarchical Clustering Method Based on Dissimilarity Matrices) analysis performed on the full set of shape variables revealed close similarities between H. moorei and both vultures. However, UPGMAs performed on PC1 and PC2 recorded separately show that H. moorei more closely resembled A. audax. This result for separate PC1 records was largely explained by the similarity in bill shape between these two large eagle species. PC2-based UPGMA highlighted differences between the two extant scavengers(V. gryphus and A. monachus), at least in part explained by phylogeny. UPGMA in PC3 showed H. moorei to be more vulture-like with respect to general skull shape, largely due to similarities in orientation of the foramen magnum and relative cranial width.
2. Finite element analysis of the skulls.- In the case of intrinsic load (bite), the mean deformation was notably lower in H. moorei than in all other taxa, although closer to A. monachus. Stress values were similar among the remaining species, except for H. sphenurus, in which stress values were high; UPGMA results were not consistent with these mean values, but showed high similarity of H. moorei with A. audax and H. morphnoides. Unlike both vultures, but like A. audax, H. moorei showed extremely low midbill stresses.
For dorsoventral loading, mean strain was intermediate in H. moorei, falling between that recorded for V. gryphus and A. monachus. Values for A. monachus were also intermediate, and values for A. audax and H. morphnoides were very low, especially for H. morphnoides. H. sphenurus presented the highest tension. These results were consistent with those of the UPGMA. The stress pattern in H. moorei was more similar to that in A. audax, with a band of high stresses across the braincase and a patch of low stresses on the side of the beak. In lateral shaking, mean stress in the skull of H. moorei was relatively high and similar to that of V. gryphus. The two extant eagle species recorded low records, although the lowest were A. monachus and the highest H. sphenurus. Under the recoil simulation, H. moorei showed relatively low stress, comparable to that of V. gryphus and A. monachus. A. audax and H. morphnoides recorded higher records, but again the highest was H. sfenuro. These results are also reflected in the UPGMA.
3. Analysis of the shape of the claws. – The analysis of the shape of the claws clearly separated the vultures from the other birds of prey. Vultures had a shorter, blunt claw with decreased curvature, and a reduced nail articular surface, whereas raptor taxa were characterized by a longer, more pointed, curved nail axis and a larger, more robust nail articular surface. UPGMA analysis performed on all claw shape variables showed that H. moorei was more like H. morphnoides.
4. Finite element analysis of the claws. – The mean stress data and the UPGMA of the stress results for the claws of the six raptor species showed that H. moorei most closely resembled A. audax, the extant eagle largest in the analysis. However, mean stress data was considerably lower in H. moorei than in any of the other raptors. H. moorei showed consistently lower stress values in the ventral region of the internal curvature, in the external curvature, and in the flexor tubercle compared to A. audax. H. morphnoides and H. sfenuro showed intermediate strain values, while the two scavengers showed the highest strain.
Once the mechanical tests, simulations and analysis have been carried out, the authors present their conclusions, which I collect below:
This work, which has not been based as the previous ones on interpretations, qualitative comparisons, as well as limited analyzes of two-dimensional forms, but on combined analyzes which have revealed a new mosaic of morphological and biomechanical characteristics attributable to the skull of H. moorei. In terms of shape, the beak is that of an eagle but its neurocranium is the closest to that of the two scavengers, specifically that of the Andean condor (V. gryphus) in particular. This contradicts interpretations based on two-dimensional analysis that the closest similarities between H. moorei and vultures were evident in the bill.
Hieraaetus moorei was also characterized by a combination of eagle and vulture traits with respect to the mechanical performance of the skull. Under intrinsic loading, mean strain in H. moorei was considerably lower than in any other taxon, but closer to that of eagles. This suggests an ability to withstand loads generated by a particularly powerful bite for its size. High mean strains recorded in V. gryphus, H. sfenuro, and H. moorei for dorsoventral pull suggest that these species are not well adapted for this behavior. Similarly, the high mean stress for H. moorei under the side-shake simulation indicates that this species was poorly adapted to perform this behavior relative to extant eagles and was again closer to the carrion-feeding V. gryphus. Interestingly, the other scavenger, A. monachus, registers a very low voltage for lateral shaking. Under recoil loading, H. Moorei and the two scavengers fared better than the remaining taxa, and Haast’s eagle was again closest to V. gryphus, which is consistent with observations of feeding and functional anatomy of the scavengers vultures.
Interpretation of both the form analysis and the FEA results of the talon show that H. moorei is unequivocally eagle-like (showing similarities to A. audax) and capable of withstanding particularly high loads. Interestingly, H. moorei showed the closest morphological similarities to the phylogenetically closest Hieraaetus: H. morphnoides. In the case of H. moorei, the increase in claw size likely reflects adaptation to killing very large prey relative to the bird’s body mass, but in this case, it could also be due to phylogenetic inertia.
In general, H. moorei’s beak is eagle-like and its braincase is vulture-like, bearing the greatest resemblance to V. gryphus between the two scavengers. Results from FEA simulations of feeding behavior show H. moorei performs very well under bite simulation, but poorly under dorsoventral and lateral loads. Under the recoil load, H. moorei outperforms the other two eagles and comes back closer to V. gryphus. Therefore, with respect to the four FEA simulations, it is closest to eagles in the ability to deliver a powerful bite, but closest to carrion-eating species such as the Andean condor. His claw, on the other hand, is like an eagle in all respects and is capable of withstanding great forces, even for his large body mass.
So how to interpret this unique combination of characters in H. moorei? Claws are the main killing tools of most raptors except falcons; however, its beak can be used to finish off a catch of prey. We propose that biting may have been more important in killing H. moorei than is typical among species of the genus Aquila. Regarding the extrinsic FEA, we first note that there is a clear difference between the two known scavengers included in the analyses. A. monachus was able to withstand high lateral loads, while H. moorei and V. gryphus performed poorly in this regard. This is consistent with the classification of A. monachus as a flesh ripper, capable of splitting open tough skin and consuming the flesh of large mammal carcasses, possibly with strong lateral forces and powerful bites. Aegypius monachus is a large, aggressive vulture that dominates other scavenging birds when fighting over a carcass occurs. In contrast, V. gryphus is a ‘swallower/swallower’ that targets the softer internal organs. Our findings of more vulture-like features in extrinsic FEAs are therefore more consistent with the proposition that H. moorei may have preferentially targeted the nutrient-rich viscera of its prey, the same behavior as most raptors that feed on carrion.
This mix of morphologies and mechanical behaviors reminiscent of obligate scavengers as well as predatory eagles may be the result of a unique combination of selective pressures. Vultures generally feed on animals much larger than themselves, but eagles usually kill and eat relatively small prey, however they can also feed on the carrion of large prey. Taphonomic evidence leaves little doubt that H. moorei killed and fed on moas, including the largest of the genus Dinornis, which could weigh more than 200 kg. Even the smallest moa species (small bush moa, Anomalopteryx didiformis; about 30 kg) was larger than Haast’s eagle H. moorei. probably inhabited open plains, as did many of its prey species. Evidently his vision wasn’t particularly sharp, but without competition, this probably wasn’t under strong selection. H. moorei used flapping flight, suggesting that flight occurred over relatively short distances. Taken together, the evidence suggests that H. moorei may have swooped down from trees or cliffs for short distances to catch and kill prey. Our results are largely consistent with the proposition that H. moorei habitually killed particularly large prey in a typically eagle-like manner using its powerful talons, but then applied feeding techniques common to vultures that eat the carcasses of species. much bigger than them. Bird skin is typically less tough than mammalian skin, so H. moorei was able to split carcasses open on its own. It is notable that H. moorei would be less adapted to eating hard tissue, such as tendons, than Aegypius, but would probably prefer viscera and muscle, like Vultur.
Our proposal that H. moorei preferred the internal organs of its prey, leading to the possibility that, in addition to vulture-like nostril scrolls and vulture-like feeding mechanics, such as most vultures had no feathers on their heads and necks. This proposition is also consistent with a Maori rock art depiction of what is believed to be an H. moorei, at Eagle Cave at Craigmore Station, South Island. The bird of prey has a dark colored body, but the head and neck remain colorless.

Bibliografía – References
García Dios, I. S. 2017. Águila calzada. Tundra Ediciones.
Gjershaug, J. O., Lerner, H. R., & Diserud, O. H. (2009). Taxonomy and distribution of the Pygmy Eagle Aquila (Hieraaetus) weiskei (Accipitriformes: Accipitridae). Zootaxa, 2326(1), 24-38.
Lerner, H. R., & Mindell, D. P. (2005). Phylogeny of eagles, Old World vultures, and other Accipitridae based on nuclear and mitochondrial DNA. Molecular phylogenetics and evolution, 37(2), 327-346.
Margalida, A., Colomer, M., Sánchez, R., Sánchez, F. J., Oria, J., & González, L. M. (2017). Behavioral evidence of hunting and foraging techniques by a top predator suggests the importance of scavenging for preadults. Ecology and evolution, 7(12), 4192-4199.
van Heteren, A. H., Wroe, S., Tsang, L. R., Mitchell, D. R., Ross, P., Ledogar, J. A., … & Sansalone, G. (2021). New Zealand’s extinct giant raptor (Hieraaetus moorei) killed like an eagle, ate like a condor. Proceedings of the Royal Society B, 288(1964), 20211913.
Comparte esto:

![]()

6 Comentarios »